
The third question in last year’s Schol Bio paper was about bats – specifically, the ecology, behaviour, and evolution of New Zealand’s only two extant native land mammals, the lesser short-tailed bat & the long-tailed bat (Mystacina tuberculata & Chalinolobus tuberculata respectively).
The long-tailed bat is a relatively new immigrant, arriving from Australia ‘just’ a million years ago or thereabouts. It’s a ‘typical’ bat, catching food (mozzies, moths & beetles) on the wing. The short-tailed bat, on the other hand, has been here at least 35 million years, with its ancestor arriving from what is now Queensland. M.tuberculata lives in dense forests and, the examiner tells us,
… are unusual bats because although they are capable of rapid, darting flight they, unlike their Australian ancestors, spend a lot of time on the forest floor searching for food. The ability to fold their wings and use them as legs allows them to scurry along the ground. Their diet consists of insects and other small invertebrates, as well as pollen, fruits and nectar of flowering plants. They are the main natural pollinator of the ground-dwelling wood rose plant. The short-tailed bat has sometimes been described ecologically as doing the job of mice.
(Remember, you can expect quite a lot of background information with some questions – make sure that you read it carefully and use it in your answers. This particular question was accompanied by quite a lot of data, from a 2006 paper by Colin O’Donnell and colleagues Jennifer Christie & Warren Simpson – and the examiner expressed some concern that many students didn’t make adequate use of all this.)
O’Donnell & his co-workers studied habitat use and nocturnal activity of the two bat species, using automatic bat detectors set up in Fiordland beech forests. These were set along transects that passed through five habitat types: red beech forest (>200m from the edge of the forest); silver beech (<100m of the forest edge); roads through the forest; on the margin between forest and grassland; and in the open grassland. Their results were presented in several graphs, some of which the examiner included – in modified form – in the exam.
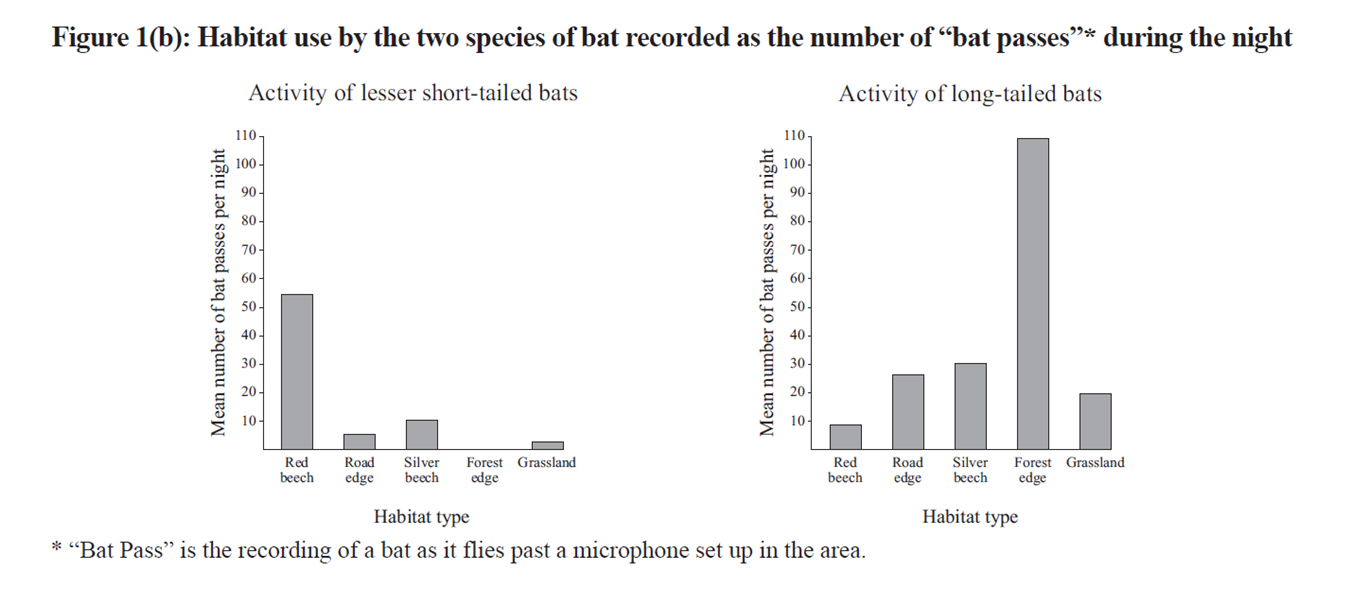
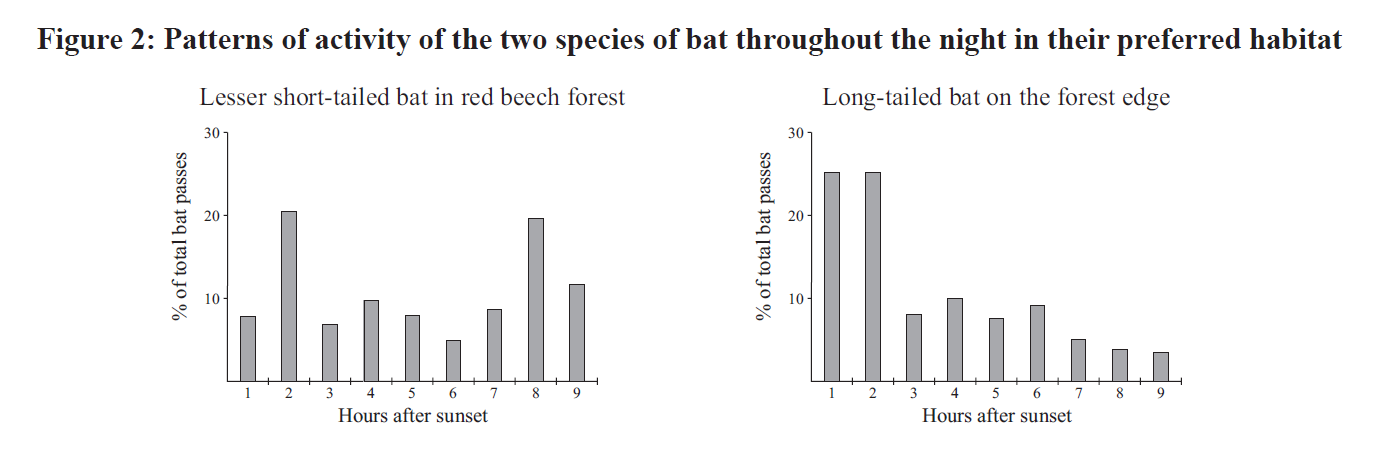
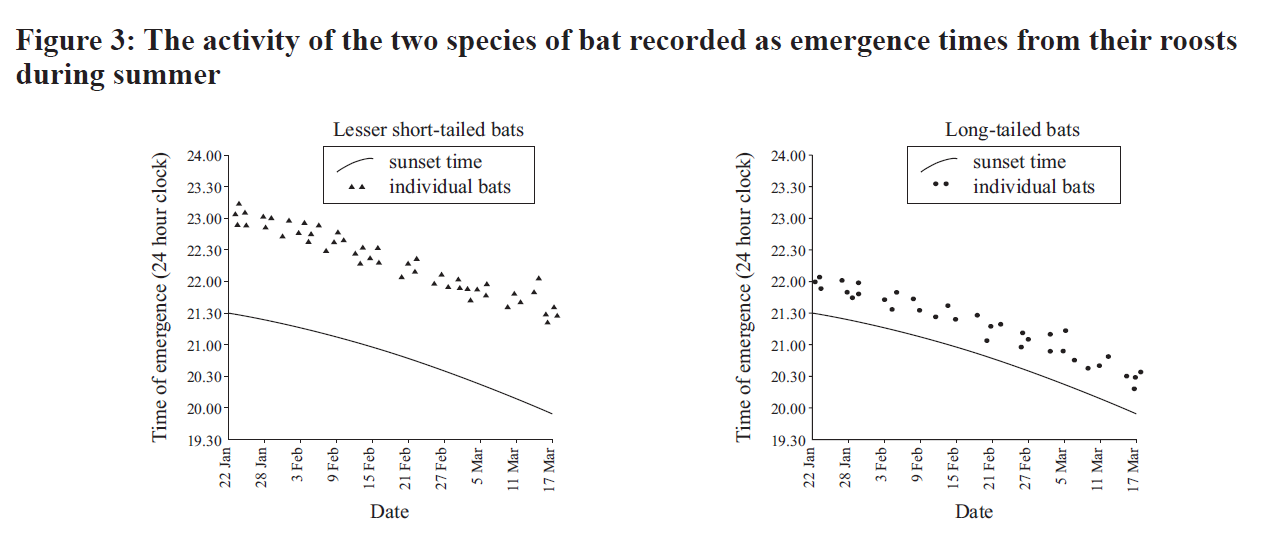
The examiner adds that "long-tailed bats were much less active in the winter months than the short-tailed bats", and then asks the question:
Discuss: the evolutionary processes that have resulted in the present-day niche of the lesser short-tailed bat in New Zealand, AND the factors that allow the co-existence of these two species of bat in New Zealand. Support your answer using data from the resource material [my emphasis – do take care to use the relevant information in your answer].
Okay – now, it’s just the short-tailed bat you’re focusing on to begin with. When that ancestral population arrived here, there’d have been no other mammalian competitors around ie vacant ecological niches. And they’d have been isolated from other bats by the Tasman sea, which sets up conditions for allopatric speciation: an isolated gene pool with no gene flow. Natural selection on advantageous mutations, plus the impact of genetic drift on this small isolated population, would have seen its gene pool diverge from that of its Australian relatives, with the potential for favourable alleles to spread rapidly. One or more of those mutations must have allowed some individuals to fold their wings to some degree in a way that allowed the animals to use them as legs when on the ground, and although these days – with so many mammalian predators around – crawling around on the ground is likely to put a small bat at a disadvantage, 30+ million years ago this wasn’t an issue. Those bats that were able to utilise this new habitat would have been able to sample a more varied diet (fallen fruit, pollen of ground-living plants), plus there would have been energy savings due to a reduction in flight: energy that could be put into growth & reproduction. The end result would be for the favourable alleles that allowed this (such as those underlying the anatomical changes allowing wing-folding) to become more common.
The relationship with the wood rose (Dactylanthus) is worth mentioning too. This is an example of co-evolution, with both members of this mutualistic relationship benefiting. The wood rose is pollinated by this little bat, which in turn gains a high-energy food supply in the form of the plant’s nectar. (The relationship is at risk due to browsing by possums, particularly in the North Island, & also to damage by rats.)
What about the co-existence of short-tailed and long-tailed bats? Well, their niches are quite different, which would reduce/minimise competition between these two species. They use their habitat in different ways (Fig. 1b): the short-tailed bat prefers beech forest while the long-tailed species is much more active along the forest margins. This alone would reduce competition for food, but (as noted in the additional information) the diets of the two bats are different. The short-tailed bat is a ground-feeding omnivore while its long-tailed cousin is an aerial insectivore. The potential for competition is reduced still further by marked differences in activity patterns ( Figs 2 & 3 above). Long-tailed bats leave their roosts to hunt about half an hour after sunset & their activity declines markedly as the night draws on, while the short-tailed species emerges 1.5 hours after the sun goes down & there’s another peak of activity near dawn ie it is ‘crepuscular’. Plus short-tailed bats are more active in winter, presumably because they’re able to make use of ground-based food sources that aren’t available to C.tuberculatus.
These differences do have implications for the conservation of lesser short-tailed bats, which are currently listed as ‘endangered’. O’Donnell et al. point out that because M.tuberculata makes little use of forest margins and open grassland, the species would not adapt successfully to fragmented habitats. This has consequences for designing reserves and managing the species – not something that you needed to comment on in answering the question, but definitely something to think about if you have an interest in conserving our native biodiversity.
[edit: I should make the point, though, that this sort of additional consideration would meet the requirement for ‘perception’ that the examiner’s looking for in recognising ‘outstanding’ scholarship performance!]
C.F.J.O’Donnell, J.E.Christie & W.Simpson (2006) Habitat use and nocturnal activity of lesser short-tailed bats (Mystacina tuberculata) in comparison with long-tailed bats (Chalinolobus tuberculatus) in temperate rainforest. NZJ Zoology 33: 113-124
steve Trewick says:
Hi
I’m marking tertiary biology scripts including a question about coevolution. Several students have said that short-tail bats and Dactylanthus provide an example of co-evolution. I did a Google search like you do and found that your Blog does say this.
This bothers me because I am not aware of scientific evidence demonstrating this evolutionary relationship. I don’t doubt the behaviour of the bats making use of wood-rose nectar, nor that this may result in pollination. But even the conclusion that this is a truly mutualistic relationship needs caution. Even if it is, mutualism and other close ecological associations, do not automatically demonstrate coevolution.
Thanks
Steve
Alison Campbell says:
Hi Steve
Apologies for the very late reply; I’ve been out of blogging for a bit. This paper provides some evidence for the statement that the relationship is coevolutionary: http://newzealandecology.org/nzje/1991.pdf
cheers
Alison