
The Level 3 & Scholarship examiners often ask you to discuss the evolutionary history of a group of organisms (Hebe, cockroaches, cicadas etc) in relation to the geological history of New Zealand. Geological changes such as the widening of the Tasman Sea, and the uplift of mountain ranges including the Kaikoura ranges & the Southern Alps, can drive evolutionary change through the isolation of populations (the Founder effect, genetic drift) & changes in selection pressures.
A new paper just out has done just this for the moa (Bunce et al., 2009), combining "mitochondrial phylogenetic information from 263 subfossil moa specimens from across NZ with morphological, ecological, and new geological data to create the first comprehensive phylogeny, taxonomy, and evolutionary timeframe for all of the species of an extinct order." This includes evidence that for much of the last 30 million years or so, the North & South islands were geographically isolated, which would have provided the basis for allopatric speciation of the fauna & flora on those 2 landmasses.
You may be reasonably familiar with the idea of adaptive radiation in relation to moa & other ratites, as it’s a common example of this pattern of evolution in senior biology textbooks in NZ. At the moment scientists think that ratites first evolved around 80 million years ago (mya) in Gondwanaland, diverging – as the supercontinent broke up – into groups that gave rise to the modern ostrich, emu, rhea, cassowary, kiwi, & the extinct elephant birds and moa. As a taxon, moa also show considerable adaptive radiation, although until recently it was difficult to tell just how many species there actually were. On the basis of morphological & aDNA data, Bunce & his colleagues suggest some changes from the till-now current taxonomy for moa, shown in the following figure (they reduce the number of species from 11 to 9, for example).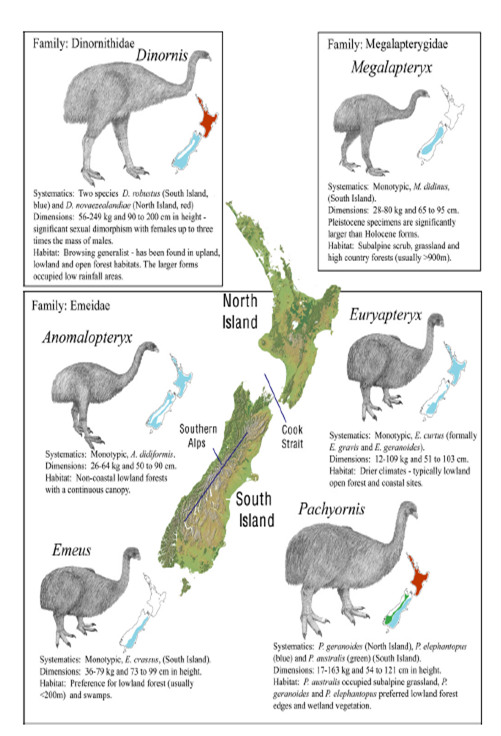
The data from ancient mitochondrial DNA also allowed the team to study the time-frame of moa evolution. Earlier studies have suggested that the moa radiation occurred about 15 mya, but Bunce et al.’s data indicate that the two main groups of moa (the Dinornithidae & Emeidae) happened just 5.27 mya – much more recently. (The 2009 paper discusses the reasons why their figure differs so significantly from the earlier one, which was also based on aDNA, & suggest this is related to how the data were calibrated.)
What was happening in NZ during that period, in ecological terms, that might have driven this divergence? One factor is the rapid orogeny (mountain-building) that we know was happening at around that time. This would have provided a range of new, untapped habitats for moa & other species to exploit – and new selection pressures as well. For example, as the Southern Alps rose up, they increasingly blocked the predominant westerly airflows, favouring the development of wet rainforests on the West Coast and a warmer, drier environment on the eastern side of the mountains, opening the way for niche specialisation and speciation. This could have been quite complex, as during glacial periods glaciers extended to the coast on the western side of the island & well down into the eastern low country, further subdividing habitats & providing additional barriers to gene flow.
In addition, from about 2 mya relatively short-lived land bridges linked the main land masses, allowing movement between the two islands. (This partly reflects the impact of glacials during the last ‘ice age’, at their peak lowering sea levels around NZ by at least 100m.)
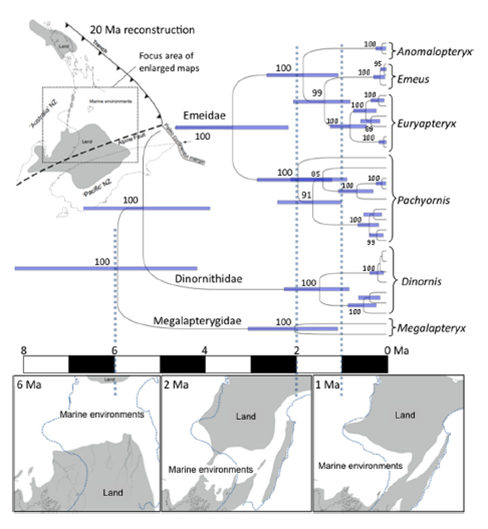
Fig.2 from Bunce et al. (2009) A spatial & temporal context for the evolution of moa. Molecular phylogeny and date estimates of the moa radiation… compared with the new paleogeographic model of Neogene New Zealand. doi/10.1073/pnas.0906660106 NB the authors caution that the timings of divergence for the various species are provisional – it would be useful to have data from many more moa specimens, to improve the accuracy of the calculations.
A great deal of new work has gone into developing those maps, particularly in the King Country, Taranaki, & central Hawkes Bay, plus data from exposed onshore marine sediments and oil exploration drill sites. During the Oligocene (> 25mya), the New Zealand land mass was reduced to a string of low, well-separated islands. Bunce et al. comment that, even when the northern landmass was re-emerging, it was still completely separated from the South Island by the Manawatu Strait (see maps above) until roughly 1.5-2 mya. Thus any land animals (& plants!) that couldn’t cross the strait were geographically isolated for that entire time. But – the moa genetic data don’t show any indication of divergence that far back, which means that the various lineages probably descended from ancestors on a single island. Fossil evidence fingers the South Island as the ancestral home, which means that moa couldn’t have colonised the North Island until less than 2 mya, only to be isolated again when Cook Strait first formed 450,000 years ago. (This interpretation is supported by data from other vertebrate remains.)
But there’s still a lot to learn! The authors conclude by saying that this "important new geological model of Neogene NZ emphasises our current lack of knowledge about the pre-Pliocene landscape, and raises important questions about the role of marine barriers and the biotic diversity of the north and south islands. The combined geological and genetic data suggests that the NZ Neogene terrestrial record is likely to have been marked by the significant loss of terrestrial endemics from a highly unusual environment, which is only just beginning to be characterised."
Bunce, M., Worthy, T., Phillips, M., Holdaway, R., Willerslev, E., Haile, J., Shapiro, B., Scofield, R., Drummond, A., Kamp, P., & Cooper, A. (2009). The evolutionary history of the extinct ratite moa and New Zealand Neogene paleogeography Proceedings of the National Academy of Sciences DOI: 10.1073/pnas.0906660106